
ニホンミツバチを飼っていると、ふとした拍子に「そもそもこの蜂は、いつから日本にいるのだろう?」と気になることがありますよね。実は近年、「ニホンミツバチは在来種ではなく、豊臣秀吉の朝鮮出兵(文禄・慶長の役、1592〜1598年)を機に朝鮮半島から持ち込まれた外来種だ」という説が、養蜂の世界でじわじわと注目を集めています。
私もこの説を初めて知ったときは、なかなか衝撃的でした。「430年前に来たばかりなら、ニホンミツバチって”日本”と名乗っていいの?」と、ちょっとモヤモヤした方も少なくないと思います。
この記事では、この「人為導入説」を頭ごなしに否定するのではなく、最新の遺伝子・ゲノム解析データと照らし合わせながら、丁寧に検証していきます。読み終わるころには、次のことが分かるはずです。
- なぜ「430年前に朝鮮から来た」という説が唱えられているのか
- 遺伝子データはその説とどう食い違うのか
- 「養蜂技術が来たこと」と「ミツバチ自体が来たこと」をどう区別すべきか
- 現時点で「在来種かどうか」にどう答えるのが一番妥当か
少し専門的な話も出てきますが、できるだけかみ砕いて説明しますので、お付き合いください。なお、この記事は生物学や歴史学の専門家による断定ではなく、公開されている研究データをもとにした整理であることを、先にお断りしておきます。

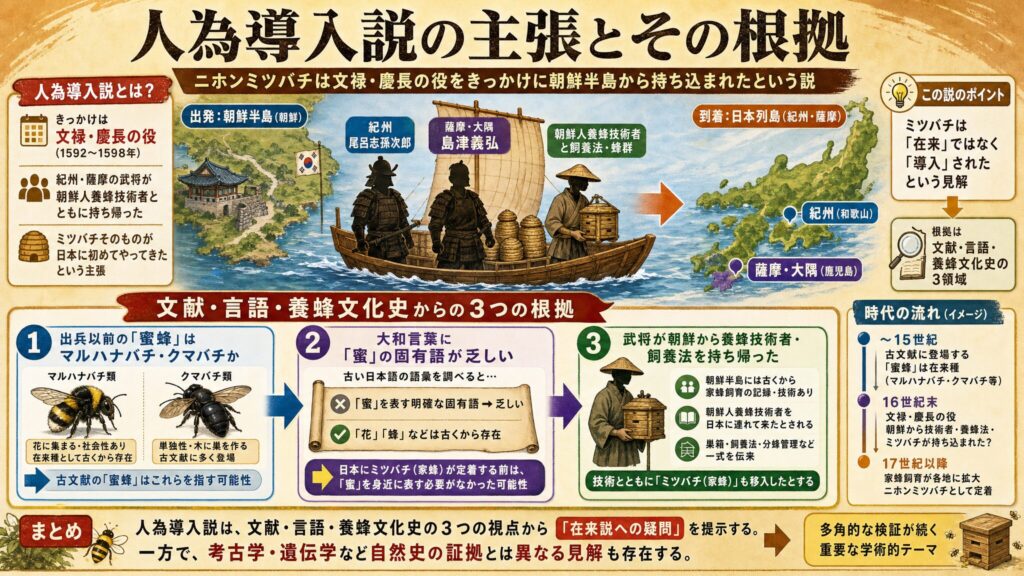
「ニホンミツバチは外来種」という説が注目されている
まずは、人為導入説がどういう主張なのかを整理しておきましょう。
文禄・慶長の役がきっかけという主張の中身
この説は、ニホンミツバチが古くからの在来昆虫ではなく、文禄・慶長の役の際に紀州の尾呂志孫次郎や、薩摩・大隅の島津義弘らによって、朝鮮人養蜂技術者とともに朝鮮半島から持ち帰られたものだ、と主張しています。代表的な論者として、東繁彦氏による『全訳 家蜂蓄養記』の解説が挙げられます(※1)。
つまり「秀吉の朝鮮出兵という具体的な歴史イベントをきっかけに、ミツバチという生き物そのものが初めて日本にやってきた」という、かなり大胆な説なのです。
文献・言語・養蜂文化史からの根拠とは
この説が拠り所にしているのは、主に文献・言語・養蜂文化史の領域です。具体的には、次のような論点が挙げられています(※1)。
- 朝鮮出兵以前の日本文献に出てくる「蜜蜂」は、現代のミツバチではなく、マルハナバチやクマバチを指していた可能性があること
- 古い大和言葉に「蜜」を意味する固有語が見当たりにくいこと
- 紀州や薩摩の武将が、朝鮮半島から養蜂技術者や飼養法を持ち帰ったとされること
たしかに、こうして並べてみると説得力があるように感じられますよね。実際、こうした文献調査の精緻さは目を見張るものがあり、養蜂文化史としては非常に重要な問題提起だと思います。
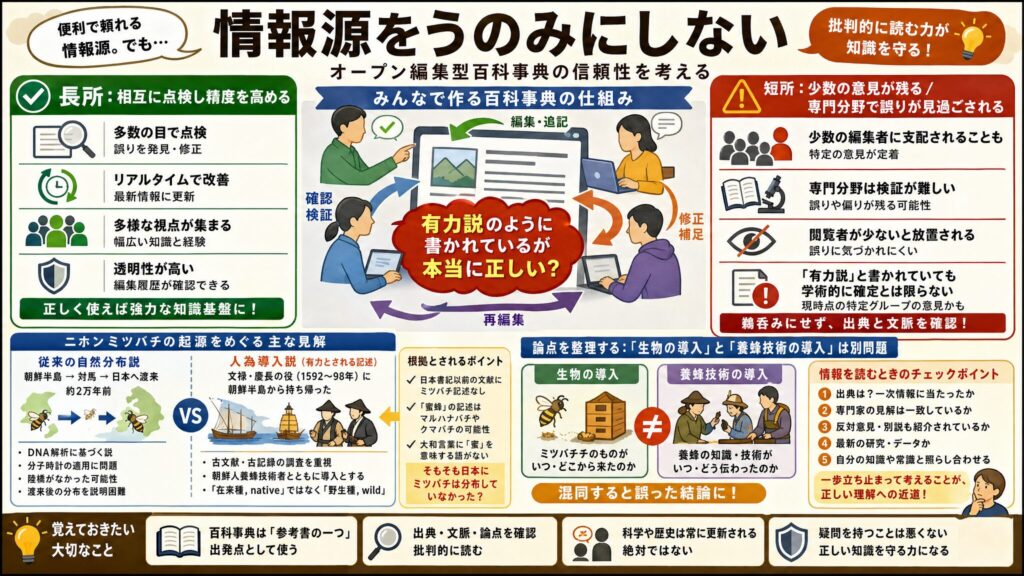
この説がなぜ広まったのか——Wikipediaは誰が書いた?
この説を信じる人が増えた理由としてWikipediaの「ニホンミツバチ」のページに以下のように記載されていることが影響していると思います。
「起源 従来、ニホンミツバチの祖先は、ミトコンドリアDNA解析から、朝鮮半島から対馬を経由して日本へ渡り、約2万年前、海面上昇による日本・対馬と朝鮮半島の分断と同時期に日本と朝鮮半島産ミツバチの遺伝的分化が生じたと考えられてきた。しかし、上の自然分布説は、分子時計仮説の適用に問題がある他、そもそも最終氷期の最寒冷期においてさえ朝鮮半島と対馬は陸橋でつながっていなかったこと、仮につながっていた場合ミツバチの南下が対馬で止まったことを合理的に説明できないことなどの問題点が指摘されている。 現在は古文献・古記録の綿密な調査分析から、ニホンミツバチの祖先は文禄・慶長の役の際に、紀州の尾呂志孫次郎や薩摩・大隅の島津義弘によって朝鮮人養蜂技術者とともに朝鮮半島から持ち帰られたものだとする人為導入説が有力に唱えられている。 日本書紀を除き、朝鮮出兵以前の文献に「蜜蜂」と記されていた生物はマルハナバチやクマバチだったことが明らかにされる一方、ミツバチについての記述が見当たらないこと、大和言葉に蜜を意味する語が存在しないことなどから、そもそも日本にミツバチは分布していなかったと考えられ、そのため現在の研究者は、家畜のセイヨウミツバチとの対比でニホンミツバチを紹介する際には「野生種, wild」という表現を用い、「在来種, native」という表現は避ける傾向にある。」(※2)
Wikipediaは、不特定多数の編集者が互いの記述を点検・修正し合うことで精度を高めていく仕組みを持っています。これは強力な長所ですが、裏を返せば、内容は「現時点で参加している特定の編集者たちの意見」にすぎず、学術的に確定した事実とは異なることがあります。特に閲覧者の少ない専門分野では、誤りや一方的な見解が見過ごされたまま残ることがあります。
ニホンミツバチの起源については、Wikipediaには特定の仮説がまるで確立した有力説であるかのような表現で記述されていると感じます。ここに書かれていることは本当に正しいのでしょうか?
一歩立ち止まって考えてみたいと思います。
論点を整理する:「生物の導入」と「養蜂技術の導入」は別問題
この議論を読み解くうえで、私が最も大切だと思うのが、「生き物そのものの導入」と「養蜂という技術・文化の導入」を分けて考えるという視点です。ここが混ざってしまうと、議論が一気にこんがらがります。
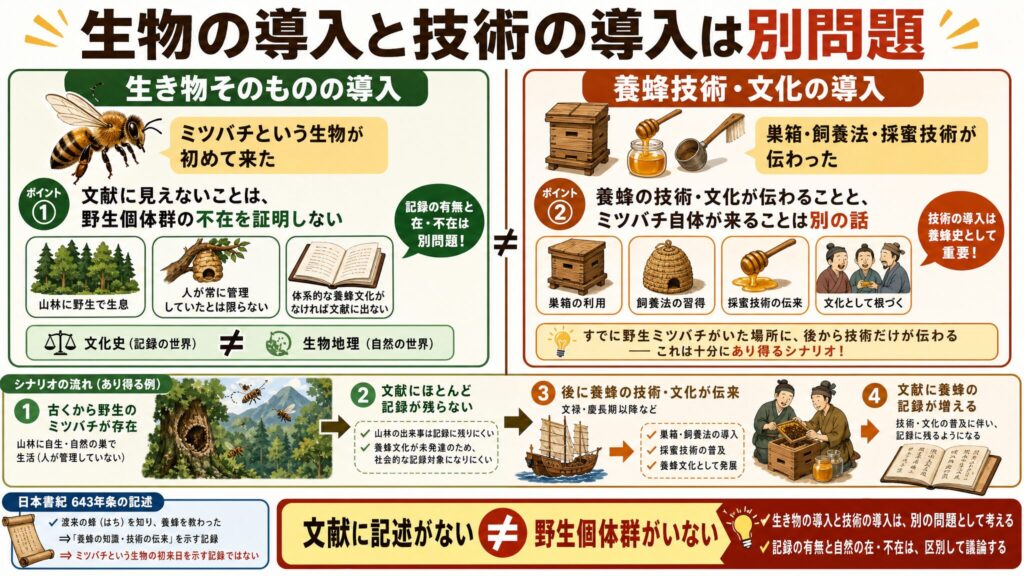
文献記録の空白は野生個体群の不在を証明しない
人為導入説の根拠の中心は「朝鮮出兵以前の文献にミツバチの記述が見当たらない」という点にあります。しかし、文献に見えないことは、野生個体群がいなかったことを直接証明するものではありません。
考えてみてください。記録に残るのは、基本的に人間社会にとって意味があった行為や産物です。ニホンミツバチのように、山林に野生の巣を作り、人間が常に管理していたとは限らない昆虫の場合、たとえ野生個体群が存在していても、体系的な養蜂文化として発達していなければ、文献にはほとんど現れない可能性があるでしょう。
つまり、文化史上の「記録の有無」と、生物地理上の「在・不在」は、きちんと区別しなければなりません。
養蜂技術が伝わることとミツバチ自体が来ることの違い
ここが核心です。文禄・慶長期以降に、朝鮮半島から養蜂技術・飼養法・巣箱利用・採蜜技術などが導入され、普及した可能性は十分にあります。これは養蜂史として重要であり、否定する必要はまったくありません。
しかし、それは「ミツバチという生物そのものが、文禄・慶長期に初めて日本列島に入った」という主張とは、まったく別の話です。すでに野生のミツバチがいたところに、後から養蜂の技術だけが伝わる——これは十分にあり得るシナリオですよね。
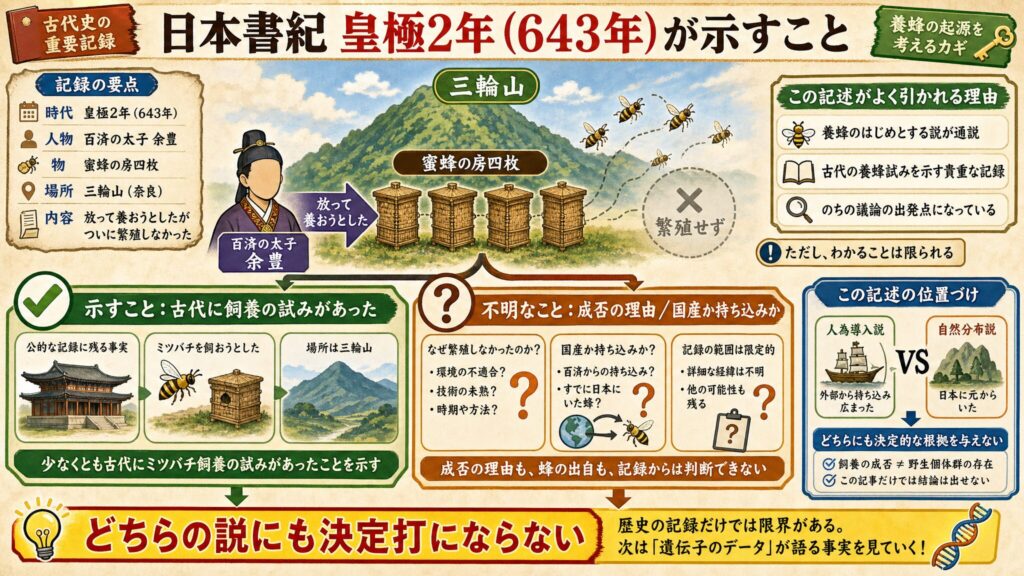
日本書紀643年条の記述は何を示すか
この文脈でよく引かれるのが、『日本書紀』皇極2年(643年)の記述です。百済の太子・余豊が「蜜蜂の房四枚」を奈良の三輪山に放って養おうとしたものの、ついに繁殖しなかった、という趣旨の記録で、これが日本における養蜂のはじめだとする説が通説になっています(※3, ※4)。
この記述は、少なくとも古代にミツバチ飼養の試みがあったことを示します。一方で、それが失敗に終わったことや、放たれた蜂が国産だったか持ち込みだったかは不明です。つまり、飼養の成否と野生個体群の存在は別問題であり、この記事は人為導入説・自然分布説のどちらにも決定的な根拠を与えないのです。
それでは、文献ではなく遺伝子に語ってもらいましょう。
短いDNA断片では、日本と韓国が「同じ」に見えてしまう
文献に限界があるなら、生き物自身が体に刻んでいる「DNAの記録」を読み解けばよい——ここで登場するのが、遺伝子データです。ただし、DNAの読み解きには一つ落とし穴があります。「日本と韓国のミツバチは遺伝的にそっくりだ」と聞いたことがある方もいるかもしれませんが、それは”DNAのどの範囲を見ているか”によって変わってくるのです。まずは、その落とし穴から見ていきましょう。
日本と韓国が同じ「Japan1型」に見える理由
短いミトコンドリアDNAの領域だけを見ると、日本・韓国のミツバチが同じ「Japan1」型に分類されることが知られています(※5)。これだけを見ると、両者はほとんど同じだと誤解されかねません。
実際、Ilyasov et al. 2018 の韓国サンプルも、非コード領域(NC2)では共通ハプロタイプ Japan1 に属し、COX1という遺伝子でも韓国・日本サンプルと同じ系統群に入りました(※5)。
見ている範囲の問題——短い断片では分からないことがある
これは、よく似た2匹の犬を見分ける場面にたとえると分かりやすいかもしれません。たとえば、目のあたりだけを切り取った写真を見ると「同じ犬だ」と思っても、顔全体を見ると別の犬だった——そんなイメージです。
短いDNA断片は「犬の顔の一部分だけを見ている状態」に近いと言えます。目のあたりが似ていても同じ犬とは限らないように、短いDNA断片が一致していても、最近の同一起源をただちに意味するわけではありません。従来の自然分布説や人為導入説の一部は、この短い断片に基づく結論を参考にしていた点に注意が必要です。
では、「顔全体」、つまり全ミトコンドリアゲノムを見ると、何が分かるのでしょうか。

全ミトコンドリアゲノムで見ると、はっきり違う——遺伝子という「時計」で430年説を検証する
短い断片では同じに見えても、DNAの「顔全体」——全ミトコンドリアゲノム——を読み解くと、話はまったく変わってきます。しかも、その違いの大きさは、両者が「いつ分かれたか」まで教えてくれます。
DNAの差が「時間」を教えてくれる——分子時計の考え方
少し基礎の話をします。生物のDNAには、世代を重ねるごとに少しずつ変異(へんい、塩基の置き換わり)が蓄積していきます。二つの集団が分かれて交わらなくなると、それぞれの集団で別々の変異がたまっていきます。
ということは、分かれてから時間が経つほど、両者のDNAの違いは大きくなるわけです。この蓄積速度をおおまかな「時計」として使い、DNAの違いの大きさから「いつ分かれたか」を推定する考え方が、分子時計です。
昆虫のミトコンドリアDNA(母から子へ受け継がれる細胞内小器官のDNA)では、二つの系統間の違いは、おおむね100万年あたり数%という速さで増えるとされます。よく使われる目安は約2.3%/百万年(※6)、別の較正では約3%台/百万年という値も示されています(※7)。
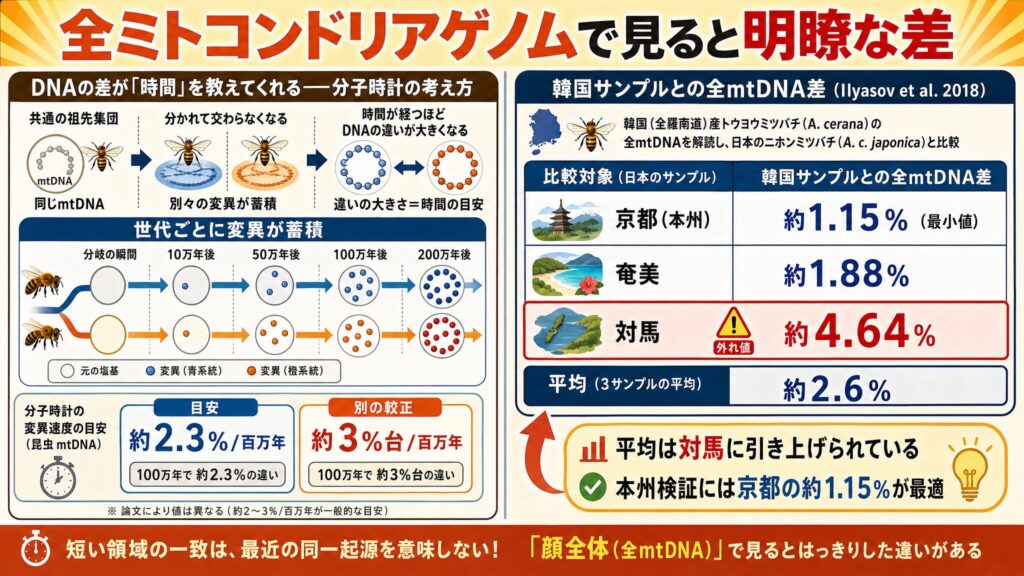
全ミトコンドリアゲノムで比べると明瞭な差が現れる
ここで重要な研究が、Ilyasov et al. 2018 です。この研究は、韓国(全羅南道)産のトウヨウミツバチ(Apis cerana)の全ミトコンドリアゲノムを解読し、日本・中国・台湾のサンプルと比較しました(※5)。
結果、韓国サンプルは日本のニホンミツバチ(A. c. japonica)と約2.58%、中国の A. c. cerana と約2.57%異なると報告されています(※5)。
ただし、この「約2.6%」という値は複数サンプルの平均で、個別の値は大きくばらつきます。論文の表から拾うと、韓国サンプルと日本各地のサンプルの差は次のようになっています(※5)。
| 比較対象 | 韓国サンプルとの差 |
|---|---|
| 京都(本州) | 約1.15% |
| 奄美 | 約1.88% |
| 対馬 | 約4.64% |
平均の約2.6%は、外れ値である対馬サンプルに大きく引き上げられたものです。ですから「文禄・慶長期に朝鮮から本州へ持ち込まれた」という説を検証するなら、本州を代表する京都サンプルとの差、すなわち約1.15%で見るのが最も適切でしょう。
注目したいのは、さきほど「Japan1型では同じに見える」と述べた韓国サンプルが、同じ個体でありながら、全ミトコンドリアゲノムではこれだけの差を抱えている、という点です。短い断片の一致は、最近の同一起源をただちに意味しない——同じ Japan1 型どうしであっても、「顔全体」で見るとはっきりした違いが隠れているのです。では、この違いはどれくらいの時間を意味するのでしょうか。
最小値1.15%でも「数十万年規模」——430年との桁違いな差
では、この1.15%を分子時計で換算するとどうなるでしょうか。
2.3%/百万年で粗く割り戻すと約50万年、最も速い側の較正を当てても約30万年規模になります(※6, ※7)。全ミトコンドリアゲノムを約16,000塩基とすると、1.15%はおよそ180塩基もの違いに相当します。
一方、もし本当に430年前に分かれたとしたら、期待される差はどのくらいでしょうか。計算すると「430年 ÷ 100万年 × 2.3% ≒ 約0.001%」、全mtDNAで見ても1塩基未満、つまり「ほぼ差がない」はずなのです。
観測された違い(約1.15%)は、430年で期待される差と桁が3つ違います。これは無視できる誤差ではありません。
分子時計の誤差を最大に見積もっても覆らない理由
分子時計に基づく推計に対する人為導入説の側からの反論として、「2.3%/百万年という速度は、もともとショウジョウバエや脊椎動物の研究から導かれ、セイヨウミツバチを経てトウヨウミツバチに適用された”仮定に仮定を重ねた”もので、科学的に厳密ではない」という指摘があります(※1)。
この指摘自体は、私も妥当だと思います。塩基置換速度は生物群によって異なり、この種の較正から精密な暦年代(何年何月という正確な年代)を出すことはできません。
しかし、ここで必要なのは精密な年代ではなく「桁」の判断です。 較正が多少ずれても、誤差はせいぜい数倍にとどまります。観測値(最小でも約1.15%)を430年に対応させるには、標準的な昆虫の置換速度を約3桁(数千倍)も速くしなければなりません。そのような較正は、どの昆虫の分類群でも報告されていないのです。
つまり、分子時計の不確かさを最大限考慮しても、観測された差は数百年ではとうてい説明できない、ということになります。

核ゲノム解析が示す日本集団の独立性
ミトコンドリアDNAは母系(母から子へ伝わる線)だけを反映します。より全体像をつかむには、両親から受け継ぐ核ゲノム(全ゲノム)の解析が欠かせません。
先ほどの犬を見分ける場面で言うと、顔つきだけでなく、平均寿命、体の大きさや体重、被毛がシングルコートかダブルコートか、毛の色、尻尾の長さといった、その犬を特徴づけるたくさんのポイントまで突き合わせるイメージです。ただし、一頭の犬を端から端まで完全に写し取るわけではありません。個体ごとに違いの出やすい「特徴的な目印」をゲノムのあちこちから多数選び出し、それらをまとめて比べる——核ゲノム解析がやっているのは、そういう作業です。
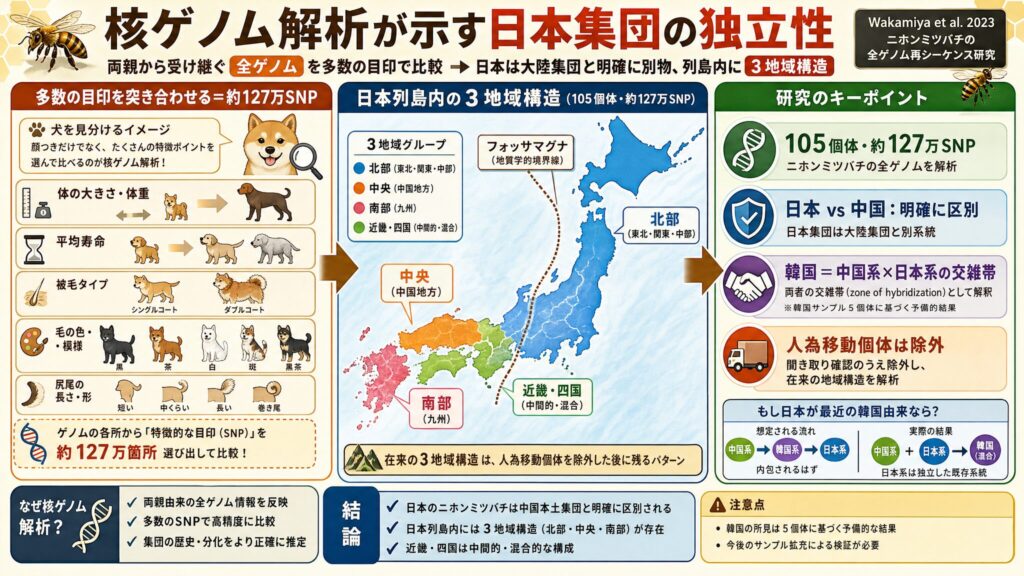
105個体・127万SNPが明かした日本列島内の3地域構造
ここで決定的に重要なのが、Wakamiya et al. 2023 の研究です。この研究は、日本列島のニホンミツバチ105個体について全ゲノム再シーケンスを行い、約127万のSNP(一塩基多型、個体間で1塩基だけ違う箇所)を検出して、列島内の遺伝構造と局所適応を調べました(※8)。
主な結果は次の二つです(※8)。
- 第一に、日本のニホンミツバチは中国本土の Apis cerana と明確に区別される。
- 第二に、日本列島内に大きく北部(東北・関東・中部)・中央(中国地方)・南部(九州)の3つの遺伝的地域集団がある。近畿・四国はそれらの中間的・混合的な構成を示す。
つまり、日本のニホンミツバチは大陸集団と明らかに別物であり、しかも列島の中でさらに3つに分かれている、ということです。
韓国集団は「中国系×日本系」の交雑帯として説明されている
特に注目したいのが、韓国に関する所見です。韓国半島のサンプルを加えた解析(補足図S4)において、韓国の個体は中国系と日本系の交雑(hybrid)として現れ、韓国半島は両者の交雑帯(zone of hybridization)と解釈されています(※8)。
これは非常に重要です。もし日本のニホンミツバチが最近の韓国由来であれば、日本系統は韓国系統に内包されるはずです。ところが実際には、逆に韓国の方が「中国系と日本系の混ざり合った地帯」として現れたのです。
つまり、日本系統は交雑の片親となる独立した既存系統であって、韓国からの新しい派生ではない、ということになります。なお、この交雑帯の所見は韓国側5個体に基づく予備的なものであり、今後の検証が必要だという点は注意が必要です。
人為移動された個体を除外した後に残る在来の地域構造
もう一つ大切な点があります。Wakamiya et al. 2023 は、近年に人為的に移動された個体(putative non-native=持ち込まれた疑いのある個体)を解析の中で特定し、飼育者への聞き取りで確認したうえで除外しています(※8)。
つまり、この研究が見ているのは、人為移動分を取り除いた後に残る「在来の3地域構造」なのです。これは、現存日本集団が「単一の最近導入された集団」として説明できるほど単純ではない、という結論を強く支持しています。
3地域集団は人為移動の証拠か、自然分化の証拠か
ここで、人為導入説の側からのWakamiya et al. 2023の解釈を検証してみましょう。「全訳 家蜂蓄養記」の第四章の最後の「補註」では、Wakamiya et al. 2023 を引用し、3地域に分かれているなら、それこそ人が運んだ証拠でであろうといった説明をしています。(※1)
「北部・中央・南部」を人為移動史と対応づける読み方
人為導入説の側に立つと、Wakamiya et al. 2023 が示した3地域集団を、文禄・慶長期以降の人為移動史の反映として読みます。たとえば、南部(九州)は薩摩の武将が朝鮮から持ち帰った蜂、北部(関東・長野)は熊野の蜂の移動、といった対応をするということになります。
たしかに、史料に出てくる「熊野」のブランド化(※4)などを思い浮かべると、もっともらしく聞こえますよね。
Wakamiya et al. 2023が示す自然分化の説明——フォッサマグナと氷期レフュジア
しかし、この解釈はWakamiya et al. 2023の論文における説明とは異なります。Wakamiya et al. 2023 自身は、3地域構造の原因を人為移動ではなく、日本列島の地質史・多様なバイオーム(生物群系)・フォッサマグナによる境界・中国地方の氷期レフュジア(避難地)・距離による隔離といった自然要因に帰しているのです(※8)。
しかも前述のとおり、Wakamiya et al. 2023は人為的に移動された個体を特定して除外したうえで、残る在来の地域構造を検出しています。つまり、3地域構造は人為移動を統制した後にもなお残るパターンなのです。
ですから「気候勾配で説明できない=人為移動の痕跡」という推論は、Wakamiya et al. 2023が自ら排除した要因を持ち込むものであり、論理の飛躍があると言わざるを得ません。整理すると、こうなります。
| 比較 | 違いの大きさ(約127万SNPの全ゲノム解析) | 著者らによる解釈 |
|---|---|---|
| 日本の集団と大陸(中国)の集団 | はっきり別のグループとして区別できる。韓国の集団は、中国系と日本系の中間的な存在 | 約10万年前に対馬海峡ができて大陸との行き来が途絶えた後、自然に分かれた(歴史時代よりはるかに古い) |
| 日本列島の中の3つの地域集団(北部・中央・南部) | 同じ解析で、互いにはっきり区別できる3グループに分かれる | 列島の成り立ち(地質)・気候や植生の多様さ・距離による自然の隔離。 |
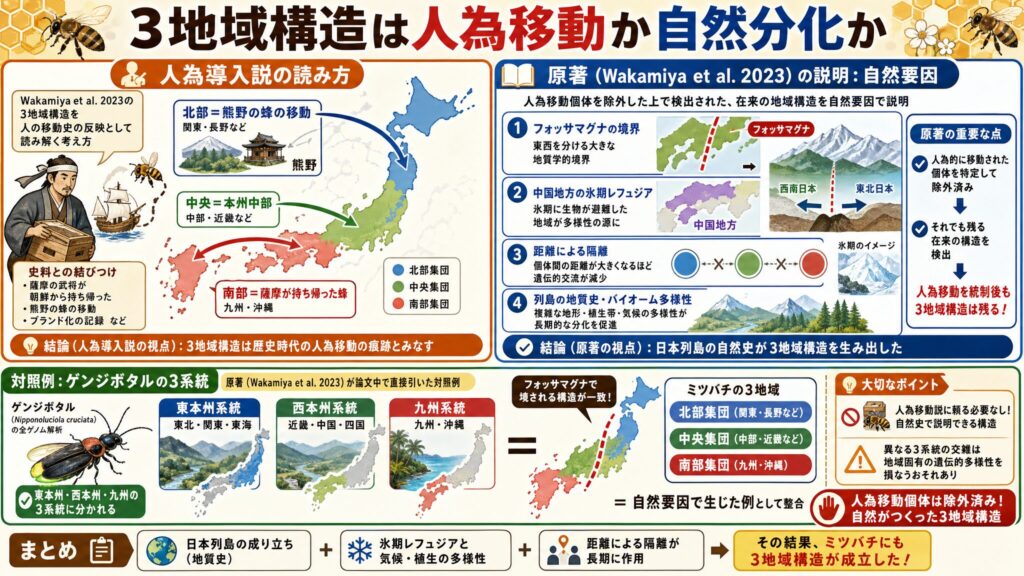
ゲンジボタルの3系統と一致する構造——原著が引く対照例
ここで面白い対照例があります。それがゲンジボタルです。
ゲンジボタル(Nipponoluciola cruciata)は日本固有の水生ホタルで、列島内に地域的な遺伝構造を持つことが知られています。重要なのは、この比較を Wakamiya et al. 2023 自身が論文中で直接引いているという点です(※8)。同論文は、ゲンジボタルの全ゲノム系統地理解析が東本州・西本州・九州の3系統を示したことを挙げ、これがニホンミツバチの北部・中央・南部の3地域構造と、いずれもフォッサマグナで境される点でほぼ一致すると述べています(※8)。
つまり、日本列島の昆虫では、同じ地質史的背景(フォッサマグナや氷期レフュジア)のもとで3地域分化が生じた可能性があるということ。ニホンミツバチの3地域構造は、その一例として理解できるわけです。
なお、最近ニホンミツバチの群れをオークションサイトで売り買いされている方がいらっしゃいますが、3地域に別れている3系統を交雑させてしまうことにもつながりますので、私としてはやめた方が良いと言う立場です。
ミツバチの起源について
ニホンミツバチが大陸のミツバチとは遺伝的に異なることがわかりましたが、ではニホンミツバチはいつから日本列島にいるのでしょうか?
まず、そもそも”祖先”にあたる大陸のトウヨウミツバチ(Apis cerana)自体が、どれほど古くから存在するのかを確認しておきましょう。
大陸のトウヨウミツバチは数百万年前からいる
ミツバチ属(Apis)はもともとアジアを起源とする古いグループで、中新世(約1600万年前ごろ)には、現在に連なるコビトミツバチ類・オオミツバチ類・洞営巣性ミツバチ類という三つの系統へと多様化していました(※9)。トウヨウミツバチはこのうち洞営巣性のグループに属し、姉妹種であるセイヨウミツバチ(Apis mellifera)と分かれたのは、約600万〜800万年前と推定されています(※10)。
かつては「約100万年前に分かれた、比較的新しい種」とみなされた時期もありましたが、その後のmtDNAやゲノムの解析によって、分岐はずっと古い数百万年スケールにさかのぼることが明らかになってきました(※10)。つまり大陸のトウヨウミツバチは、人類が登場するよりはるか以前から、東アジアに広く暮らしてきた古い野生種なのです。
では、その古いミツバチが日本列島にいた証拠はあるのでしょうか。実はごく最近、それを示す化石が見つかりました。
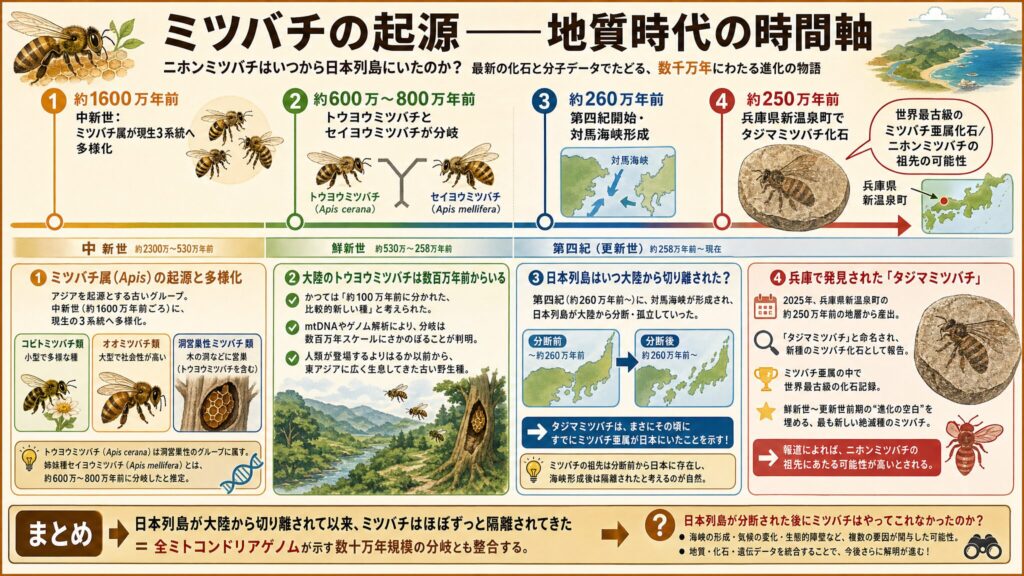
兵庫で発見されたタジマミツバチ——世界最古級のミツバチ亜属化石
2025年、京都産業大学と慶應義塾幼稚舎の研究チームが、兵庫県新温泉町の約250万年前の地層から産出した化石を「タジマミツバチ」と命名し、新種のミツバチ化石として報告しました(※11, ※12)。
このタジマミツバチは、現在養蜂に使われるミツバチなどが属する「ミツバチ亜属」の中で最も古く、これまで化石記録がなかった鮮新世〜更新世前期の時代に生息していたとされます。世界最古のミツバチ亜属の化石であると同時に、最も新しい絶滅種のミツバチでもあるという、進化の空白を埋める発見です(※11)。
報道によれば、このタジマミツバチはニホンミツバチの祖先にあたる可能性があるとされています(※13)。地球が氷河期に入り始めた頃、すでに日本列島にミツバチ亜属が生息していた——つまり、数百万年前から大陸にいたミツバチの仲間が、遅くとも約250万年前には日本にもいた、ということです。
対馬海峡が形成され、日本列島が大陸から切り離されていったのは第四紀(約260万年前〜)とされますが、タジマミツバチは、まさにその頃に、すでにミツバチ亜属が日本にいたことを示しています(※11)。
つまり、ミツバチの祖先は約260万年に日本列島が大陸から分断される前から日本列島にいて、海峡ができた後には隔離された——と考えるのが自然です。これは、全ミトコンドリアゲノムが示す数十万年規模の分岐とも、きれいに整合します。列島が孤立して以来ほぼずっと隔離されてきたからこそ、それだけの差が蓄積したわけです(※5)。
日本列島が大陸から切り離された後にミツバチはやってこれなかったのか
大陸や日本列島に数百万年も前から暮らしてきたミツバチは、日本列島が大陸から分断された後には全く渡って来ることはなかったのでしょうか?
実はこの「大陸と日本がいつ・どうつながっていたか」という問題、特に南方(対馬)ルートでは、近年の複数の可能性が指摘されているようです。最新の議論を踏まえて、整理してみます。
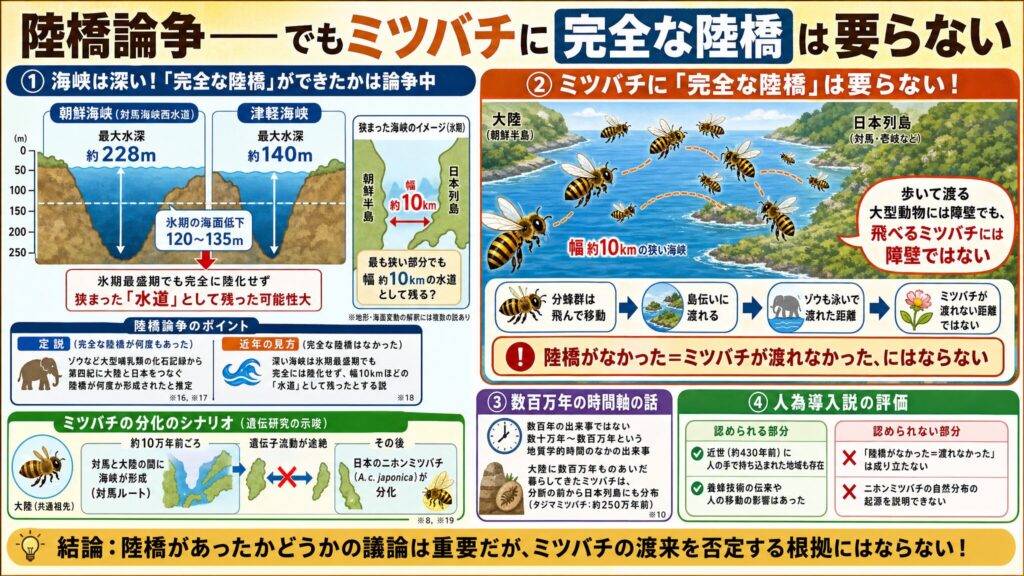
深い海峡——「完全な陸橋」があったかは論争中
日本列島と大陸の間にある海峡は、意外に深いものです。対馬と朝鮮半島の間の朝鮮海峡(対馬海峡西水道)は最大水深が約228メートルと深く(※14)、最も浅い按部でも100メートルを超えます。北海道と本州の間の津軽海峡も約140メートルあります。一方、氷期の海面低下はおおむね120〜135メートル(※15)。海峡の深さとほぼ同じか浅い程度なので、「氷期に大陸と陸続きの完全な陸橋ができたかどうか」は、専門家の間で議論があるようです。
定説は、哺乳類の化石記録を根拠にしています。ゾウの仲間など大型哺乳類が大陸から渡来した形跡から、第四紀の何度かの時期に大陸と日本をつなぐ連絡が生じた、と推定するものです(※16, ※17)。
一方、近年、これらの深い海峡は氷期最盛期でも完全には陸化せず、最終氷期でも朝鮮海峡は幅10キロメートルほどに狭まった「水道」として残っていた、とする見方が提唱されています。堤(2021)は、第四紀(約260万年前〜)に入って以降、本州が大陸と完全に地続きになった時期はなかった、という立場を示しています(※18)。つまり「ゾウが歩いて渡れる完全な陸橋が何度もあった」という見方に対して違う見方が指摘されているのです。
一方で、ニホンミツバチの遺伝研究において、Wakamiya et al. 2023 は、対馬ルートを最有力とし、約10万年前に対馬と大陸の間の海峡が形成されて遺伝子流動が途絶えた後に、日本のニホンミツバチ(A. c. japonica)が分化した、というシナリオを採っています(※8, ※19)。
でも、ミツバチに「完全な陸橋」は要らない
ここで大切なのは、この陸橋論争が、ミツバチの渡来にはほとんど影響しないという点です。なぜなら、ミツバチは陸橋を必要としないからです。分蜂群は飛んで移動し、壱岐や対馬を島伝いに、狭まった海峡を越えていくことができます。氷期に幅10キロメートルほどに狭まった海峡は、飛べるミツバチにとって、越えられない障壁ではありません。象さえも泳いで渡れた距離ですしミツバチに渡れなかったとは思えません。
ここで、冒頭に引用したWikipediaの一節を思い出してください。そこには「最終氷期の最寒冷期でさえ朝鮮半島と対馬は陸橋でつながっていなかった」とありました(※2)。堤説のみを事実として記載しているわけですが、これが仮に事実であったとしても、それは、歩いて渡るしかない大型動物が渡れなかったことを意味するだけで、飛んで渡れるミツバチまで渡れなかったことにはなりません。つまり「陸橋がなかった」としても、「ミツバチが430年前に人の手で持ち込まれた」という主張の証拠にはならないのです。
数百年ではなく、地質学的な時間軸の出来事
「完全な陸橋があったか」には議論がありますが、ミツバチに関するかぎり、その答えは結論を左右しません。
大陸に数百万年ものあいだ暮らしてきたミツバチ(※10)は、約250万年前のタジマミツバチが示すように、日本列島が大陸から分断される前から日本列島にも分布していた。
そして、日本列島が分断した後にも、氷期に狭まった海峡が狭まった時期には大陸から日本列島にやって来ることができた。
数百年前の出来事ではなく、数十万年から数百万年という地質学的な時間軸のなかの出来事と考える方が妥当と思います。
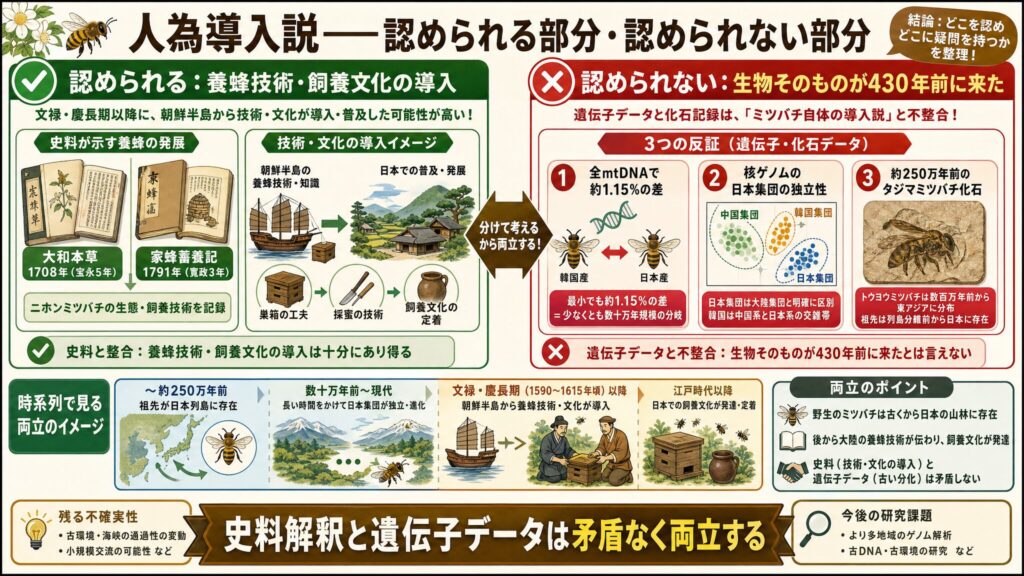
人為導入説で認められる部分・認められない部分
ここまでの議論を踏まえて、人為導入説を全否定するのではなく、どこを認め、どこに疑問を持つべきかを整理してみます。
養蜂技術・飼養文化の導入はあり得る
まず、認めるべき部分です。文禄・慶長期以降に、朝鮮半島から養蜂技術や飼養文化が導入・普及した可能性は十分にあります。これは養蜂史として重要であり、否定する必要はありません。
実際、日本の養蜂が本格化したのは江戸時代からで、宝永5年(1708年)の『大和本草』や寛政3年(1791年)の『家蜂蓄養記』など、ニホンミツバチの生態や飼養技術を述べた書物が次々に登場しています(※4)。技術や文化が大陸由来の影響を受けながら発展していったこと自体は、十分にあり得る話です。
「ミツバチ自体が430年前に来た」とは言えない理由
一方で、認められない部分があります。それは「ミツバチという生物そのものが、文禄・慶長期に初めて日本列島に入った」という主張です。
ここまで見てきたとおり、遺伝子データはこの主張と整合しません。
- 全ミトコンドリアゲノムでは、韓国産と日本産の間に最小でも約1.15%の差があり、これは少なくとも数十万年規模の分岐を意味する(※5)
- 核ゲノム解析でも、日本集団は大陸集団と明確に区別され、韓国はむしろ中国系と日本系の交雑帯として現れる(※8)
- そもそも大陸のトウヨウミツバチは数百万年前から東アジアに分布しており(※10)、約250万年前のタジマミツバチ化石が示すように、列島が大陸から切り離される頃には祖先がすでに日本にいたと考えられる(※11)
史料解釈と遺伝子データは矛盾しない形で共存できる
大切なのは、この二つは矛盾なく両立する、ということです。
野生のミツバチがすでに山林にいたところへ、後から大陸の養蜂技術が伝わり、飼養文化が発達した——こう考えれば、史料の示すこと(技術・文化の導入)と、遺伝子データの示すこと(生物の古い分化)は、何も衝突しないのです。
つまり「養蜂という技術・文化の導入」と「ミツバチという生物そのものの導入」を分けて考えれば、人為導入説の文化史的な価値を尊重しつつ、生物学的な主張だけを退ける、という公正な整理ができるわけですね。
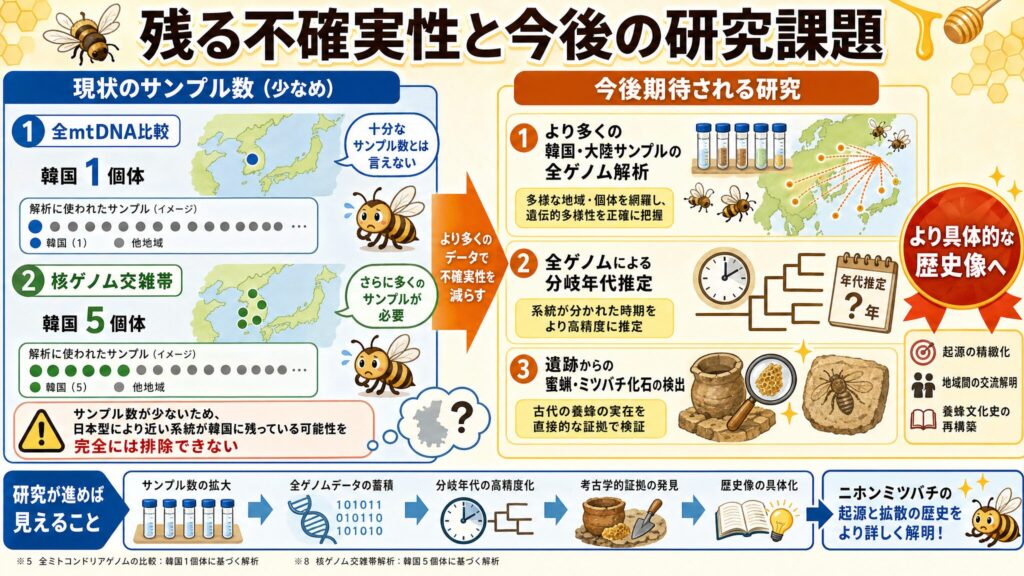
残る不確実性と今後の研究課題
ここまでかなり踏み込んで書いてきましたが、残る不確実性もきちんと挙げておきます。「これで完全決着」と断言するつもりはありません。
韓国側サンプル数が少な目
少し気になったのは韓国側の蜜蜂サンプル数が少な目であることです。全ミトコンドリアゲノムの比較は韓国1個体(※5)、核ゲノムの交雑帯解析は韓国5個体(※8)に基づいています。十分にサンプリングされていない地域に、日本型により近い系統が残っている可能性は、完全には排除できません。今後、より多くの韓国・大陸サンプルの全ゲノム解析が行われるとこのような議論を排除できると思います。
さらに研究が進めば見えること
今後への期待です。
もし、今後より多くのサンプルを用いた全ゲノムデータによる分岐年代推定、遺跡からの蜜蝋(みつろう)やミツバチの化石の検出などが更に進めば、より具体的な歴史像が描けるようになるでしょう。
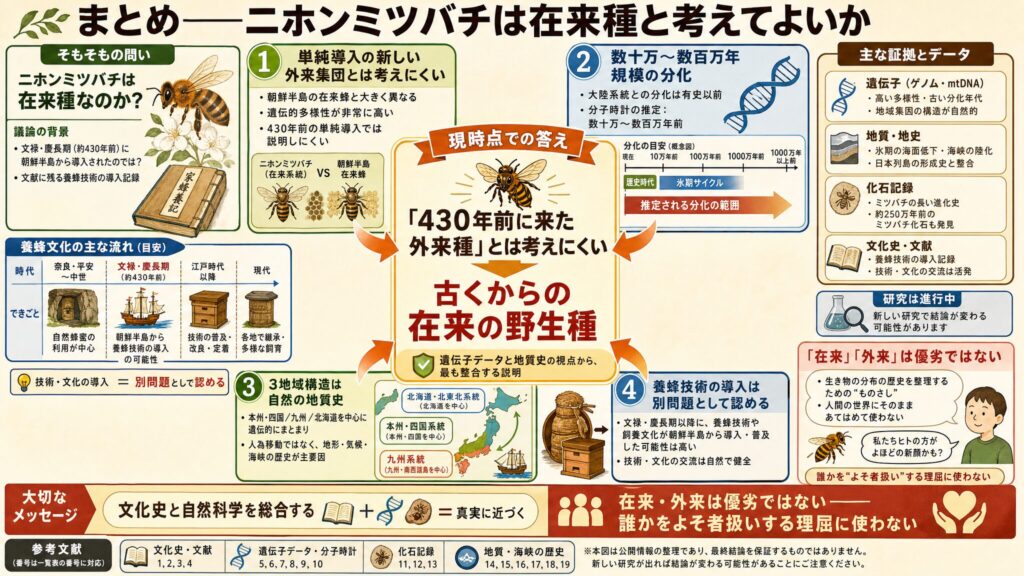
まとめ:ニホンミツバチは在来種と考えてよいか
長くなりましたので、最後に全体を整理しましょう。
遺伝子データが示す「最も妥当な整理」
現時点で最も妥当な整理は、次のようになると思います。
- 現存するニホンミツバチの主要系統は、文禄・慶長期に朝鮮半島から単純に導入されて成立した「新しい外来集団」とは考えにくい
- 遺伝子データは、少なくとも歴史時代よりはるかに古い分化(数十万〜数百万年規模)を示している(※6, ※7, ※5, ※8)
- 列島内の3地域構造の主たる成因も、人為移動ではなく自然の地質史と考えられる(※8)
- 一方で、養蜂技術や飼養文化が文禄・慶長期以降に朝鮮半島から導入・普及した可能性は、別問題として認められる(※4)
「在来種かどうか」という問いへの現時点での答え
では、冒頭の問い「ニホンミツバチは在来種なのか?」に、現時点でどう答えるべきでしょうか。
私の理解では、こうなります。「430年前に朝鮮から来たばかりの外来種」とは、遺伝子データを解析した論文からみて考えにくいです。 少なくとも有史時代をはるかにさかのぼる時期に大陸系統と分化し、列島内で自然に地域分化してきた、古くからの「在来の野生種」と考えるのが、現在のデータと最も整合します。
ニホンミツバチをめぐる科学と文化史の面白さ
私がこのテーマで一番面白いと感じるのは、文献・言語という文化史のアプローチと、ゲノム・化石という自然科学のアプローチが、互いに違う角度から同じ生き物の起源を検証しているという点です。
人為導入説は、養蜂文化史への貴重な問題提起として大きな意味があり、特に「全訳 家蜂蓄養記」は労作と考えます。同時に、遺伝子データは「生き物としてのニホンミツバチ」の古さを語っています。
この二つを対立させるのではなく、それぞれの知見を総合的に組み合わせて更に真実に近づきたいですね。
なお、この記事は公開されている研究データをもとにした整理であり、生物学的・歴史学的な最終結論を保証するものではありません。新しい研究が出れば結論が変わる可能性があることにご注意ください。
最後に——「在来」「外来」について
「在来種」「外来種」は、生き物の分布の歴史を整理するための生態学のものさしであって、どちらが「えらい」とか「本物」とかを決める言葉ではありませんし、まして人間の世界にスライドしてそのまま持ち込んでよいものではありません。
そもそも、数十万年前以上前からこの列島で暮らしてきたニホンミツバチからすれば、数万年前に来た私たちヒトの方がよほどの新顔。「最近わざわざ巣箱まで用意してくれる、あの大きな外来種」とでも思われているかもしれません。この記事を誰かを”よそ者”扱いする理屈には使わないようにお願いします。
参考文献
ニホンミツバチの起源に関心を持たれた方は、ぜひ下記の参考文献にあたってみてください。
- 久世松菴(著), 東繁彦(訳注・解説)『全訳 家蜂蓄養記—古典に学ぶニホンミツバチ養蜂』農山漁村文化協会、2023年
- 「ニホンミツバチ」Wikipedia (2026年6月9日時点)— https://ja.wikipedia.org/wiki/%E3%83%8B%E3%83%9B%E3%83%B3%E3%83%9F%E3%83%84%E3%83%90%E3%83%81
- 「みつばちの意外なお話 日本の養蜂ことはじめ」山田養蜂場 — https://honey.3838.com/surprise/kotohajime.html
- 「養蜂の歴史」一般社団法人日本養蜂協会 — https://beekeeping.or.jp/beekeeping/history/
- Ilyasov, R. A., Park, J., Takahashi, J., & Kwon, H. W. (2018). Journal of Apicultural Science, 62(2), 189–214.(韓国産 Apis cerana の系統学的独自性)
- Brower, A. V. Z. (1994). PNAS, 91, 6491–6495.(昆虫mtDNA分子時計の較正)
- Papadopoulou, A., Anastasiou, I., & Vogler, A. P. (2010). Molecular Biology and Evolution, 27(7), 1659–1672.(昆虫mtDNA分子時計の再検討)
- Wakamiya, T. et al.「Genetic differentiation and local adaptation of the Japanese honeybee, Apis cerana japonica」Ecology and Evolution, 13(10), e10573 (2023) — https://doi.org/10.1002/ece3.10573
- ミツバチ属はアジアを起源とし、中新世(約1600万年前ごろ)に現生の3系統(コビトミツバチ類・オオミツバチ類・洞営巣性ミツバチ類)へ多様化したとされる(Raffiudin & Crozier 2007; Engel 1999 ほか)
- トウヨウミツバチとセイヨウミツバチの分岐年代は約600万〜800万年前と推定されている(Garnery et al. 1991; Sheppard & Berlocher 1989 ほか。近年のゲノム解析では約600万〜2500万年前と幅をもつ推定もある)
- 「約250万年前の地層からミツバチの新種化石『タジマミツバチ』を発見」京都産業大学 — https://www.kyoto-su.ac.jp/news/news-001874.html
- 「プレスリリース 慶應義塾」 — https://www.keio.ac.jp/ja/press-releases/2025/10/27/28-170262/
- 「250万年前の化石からミツバチ、新種と特定 慶応義塾幼稚舎など」日本経済新聞 — https://www.nikkei.com/article/DGXZQOSG3072A0Q5A031C2000000/
- 「朝鮮海峡」Wikipedia(最大水深約228mは海上保安庁海洋情報部の海図に基づく)— https://ja.wikipedia.org/wiki/朝鮮海峡
- 氷期の海面低下と日本海周辺の海峡の陸化について。「URBAN KUBOTA」No.31 ほか。なお、海峡の地史の一次資料としては大島和雄「最終氷期以降の日本周辺海峡の地史」第四紀研究 29, 193–208 (1990) 等があり、Wakamiya et al. 2023 もこれを引用しています
- 樽野博幸「哺乳類化石の変遷から見た日本列島と大陸間の陸橋の形成時期」第四紀研究, 49(5), 309–314 (2010) — https://www.jstage.jst.go.jp/article/jaqua/49/5/49_5_309/_article/-char/ja/
- 河村善也「第四紀における日本列島への哺乳類の移動」第四紀研究, 37(3), 251–265 (1998) — https://www.jstage.jst.go.jp/article/jaqua1957/37/3/37_3_251/_article/-char/ja/
- 堤之恭『新版 絵でわかる日本列島の誕生』講談社、2021年(第四紀以降、本州が大陸と完全に地続きになった時期はないとする立場)
- 「対馬について」対馬野生生物保護センター — https://torayama-twcc.jp/tsushima/









コメント